
Pamela A. Davol, 76 Mildred Avenue, Swansea,
MA 02777-1620.
pdavol@labbies.com
B/b, E/e, and Beyond:
A Detailed Examination of Coat Color Genetics
in the Labrador Retriever
Why do yellow Labs have variations of shading? What causes the fox-red color in yellow Labs? Why do some Labs have white spots on their chests? What causes some Labs to have white at the base of the hair shafts on their tails? Why do many Labs develop gray hairs as they get older? The answers to these questions and many others pertaining to coat color can be found in the canine genes that influence production of pigment, also known as melanin. These same genes determine the black, chocolate or yellow coat color found in the Labrador breed, however, to address these former questions, one must explore beyond the genes responsible for simple solid coloration. The following article presents a detailed examination into the factors responsible for inheritance of coat color including an explanation of the modifying genes responsible for unique coat-color attributes.
Table of Contents
DNA... Chromosomes... Genes: The Ancestral Legacy
In 1944, experiments conducted by Oswald Avery and colleagues identified DNA (deoxyribonucleic acid) as the material in the cell responsible for carrying genetic information. Thereafter, DNA was aptly referred to as "the thread of life." The sequences of nucleotides, small molecules linked together that form two complimentary chains to compose the native double-helix form of the DNA molecule, are responsible for encoding specific proteins required for many cellular functions. These proteins are critical for proper function for organs that, in turn, are required for the normal function and life of the whole organism.
In canine somatic cells-- cells responsible for composing the body--, the DNA is packaged into 78 chromosomes. Thirty-nine of the chromosomes contain DNA passed on, or inherited, from the sire's germ cell (sperm) and the other 39 contain DNA inherited from the dam's germ cell (egg). Each chromosome, therefore, has a partner-chromosome inherited from the other parent so that the 78 chromosomes form 39 pairs. Germ cells differ from somatic cells in that they have divided through a process known as meiosis. As a result of meiosis, each germ cell receives one of the partner-chromosomes so that it has one chromosome of each type. Meiosis, therefore, provides the means by which offspring may inherit genetic information from each parent, while maintaining the same number of chromosomes in its somatic cells as found in the somatic cells of each parent and in other individuals within its species.
When germ cells fuse during fertilization they produce a single somatic cell from which the offspring will develop. This somatic cell will divide to produce more cells by the process of mitosis. During mitosis, before a somatic cell divides, each chromosome will be duplicated so that a canine cell just prior to dividing will have 156 chromosomes (or 78 chromosome pairs). When the somatic cell divides, each new cell will have the same amount of genetic information as the former cell. Mitosis, therefore, provides the means for a single fertilized cell to develop into a whole organism, while maintaining the same chromosome number and amount of genetic information in each cell throughout the body.
Composing each of the chromosomes, the DNA for encoding specific proteins that are responsible for producing individual genetic traits, or characteristics, are called genes. Each gene encoding a specific protein is located at a particular site on the chromosome called a gene locus. The location of the gene locus for a trait on one chromosome will match the location of the gene locus for that same trait on its partner-chromosome. However, although the location of a gene locus is identical for each partner-chromosome, the nucleotide sequence of the gene may be different on each of the two paired-chromosomes. When this occurs and the nucleotide sequence of the DNA to encode a specific protein is altered, a condition referred to as a mutation, the resulting protein may physically and functionally differ from the normal protein. For this reason, because more than one nucleotide sequence of a gene may be present at a gene locus on partner-chromosomes, alternate sequences controlling one particular trait are referred to as alleles.
Offspring may inherit the same allele at a particular gene locus from each parent. In such an instance, the offspring is considered homozygous for the particular trait governed by that gene locus. Alternatively, offspring may inherit one allele from one parent but a different allele from the other parent. In this instance, the offspring is considered heterozygous for the particular trait. When offspring are heterozygous at a particular gene locus, often the characteristic of only one of the alleles will be expressed by the offspring. In such an instance, the allele that results in expression of the trait is considered dominant over the other allele which is considered recessive. A simple example of dominant and recessive alleles, respectively, in Labrador coat color genetics are the "B" allele for expression of black and the "b" allele for expression of chocolate. Black Labradors may be homozygous for black (BB) or heterozygous (Bb). In the latter instance, the dog carries an allele for the chocolate coloration, but its expression is masked by the dominant allele for black. Heterozygous Labs may produce chocolate offspring only if they are bred to another heterozygous Lab or a Lab homozygous for the recessive "b" (i.e. a chocolate Lab). For a Labrador to appear chocolate, it must inherit the "b" allele from both parents.
The Basis of Dominant and Recessive Alleles
Though many individuals are familiar with the terms dominant and recessive, some, if not most of these individuals are unaware of the underlying mechanisms that make some alleles dominant and others recessive. To understand these mechanisms, one must go back to the DNA sequence of the gene.
The DNA of each gene locus is responsible for producing a single protein. As mentioned earlier, DNA is composed of molecules called nucleotides. When these nucleotides are arranged in specific order they encode for amino acids, which are molecules that make up proteins. The sequence of amino acids, in turn, will determine protein structure and function. Therefore, for normal cellular function, the correct sequence of DNA all the way through to the sequence of amino acids must be conserved. When mutations occur in the DNA nucleotide sequence, the resulting amino acid sequence may be altered. As a result, the protein for that gene locus will either not be produced, or will be different from the normal protein. In the latter instance, the alternate protein may function adequately but produce some physiological changes not seen in the presence of the normal protein. Therefore, different alleles occur as a result of mutations in the DNA for a particular gene locus.
In many cases, recessive alleles are those that produce no protein. In other cases or in cases where there are more than two possible alleles for a gene locus (discussed below) usually there is a rank order of dominance. In such an instance, the DNA mutation results in a structural change in the protein that influences its reactivity. Put more simply, those alleles that produce proteins with the greater capacity to function will be more dominant to alleles that produce weakly-functional or non-functional proteins. Therefore, in the case of a heterozygous (Bb) black Lab, the black producing allele produces a protein that functions more efficiently than the protein encoded by the chocolate producing allele.
Melanin: one molecule, many shades of coat color
Despite the fact that the B locus on a chromosome may be occupied by either the "B" or the "b" allele, the same molecule responsible for the black coloration in the Lab is also responsible for the chocolate coloration. Melanin, or pigment molecules, are produced by and packaged into small organelles called melanosomes. Within the melanosomes are enzymes called tyrosinase, which are proteins needed to make melanin from molecules called tyrosine, and structural matrixes upon which the melanin is organized after it is made. The actual color of the melanosomes is determined by the amount of melanin it contains. Melanosomes are produced by specialized cells called melanocytes. Melanocytes are distributed throughout various areas of the body including the eyes, the hair, and the skin where they transfer the melanosomes into the cells that compose these structures. Color of each structure will be determined by both the color of the melanosomes it contains as well as the distribution of the melanosomes within its cells.
In hair comprising the coat of dogs, two types of melanin have been found. Eumelanin is responsible for black/brown pigment, and phaeomelanin is responsible for red/yellow pigment. Phaeomelanin is formed by a modification of the pathway that leads to production of eumelanin from tyrosine. It is important to know that phaeomelanin has only been found in cells composing hair and not in cells composing the skin. In Labs, eumelanin and phaeomelanin production are controlled by the E locus. The protein product of other gene loci will determine the level of pigment expression. For example, in the instance of black versus chocolate coat color, color appearance is actually determined by the B locus. The B locus controls pigment not by controlling changes in the actual eumelanin molecule, but rather by controlling number, size, and pattern of the distribution of melanosomes in the hair shafts. Therefore, Labs that are homozygous for black at the B locus have large eumelanin-producing melanosomes that are evenly and densely packed throughout the hair shaft. Labs that are homozygous for chocolate have melanomsomes that are less tightly packed into the hair shaft.
The Function of the E Locus and the A Locus in the Labrador Retriever
The E gene locus in Labs determines whether the dog will be black/brown (eumelanin) or red/yellow (phaeomelanin). This locus encodes the melanocyte stimulating hormone receptor (a.k.a. the melanocortin 1 receptor; Mc1r). Labs that are homozygous for the dominant E allele have a constitutively active, mutant form of Mc1r; that is, the receptor is always "turned-on", even in the absence of melanocyte stimulating hormone (MSH). As such, eumelanin is constantly produced and the dog appears black or chocolate. Labs that are homozygous for the recessive "e" allele also have a mutant form of Mc1r. This mutant, however, is a "loss of function" receptor that cannot produce eumelanin, even in the presence of MSH. Therefore, Labs that are homozygous for the "e" allele can only produce phaeomelanin and, therefore, will appear yellow.
In some other breeds, the Agouti locus is responsible for determining yellow color. Some of the recessive agouti alleles produce molecules that inhibit the activation of Mc1r by interfering with binding of MSH to the receptor. In these dogs, Mc1r is a normal receptor type (called wild-type; E+); that is, it will only produce eumelanin when activated by MSH and will only produce phaeomelanin when the receptor is inhibited. As such, these breeds often display a combination of black as well as tan (yellow) coloring resulting from production of both eumelanin and phaeomelanin. The Agouti genes are often ignored in the Labrador with most writers stating that all Labs are As at the agouti locus. This information is based solely on the observation that agouti will cause both black and tan banding of the hair shaft and since the black banding is not present in Labs, Labs must be As (the allele that encodes the agouti suppressor). This argument, however, fails to take into consideration the presence of the mutant "loss of function" receptor in yellow Labs compared to these other breeds. The Mc1r in yellow Labs is unable to produce eumelanin pigment under any circumstances. Instead, the effects of the recessive Agouti alleles in yellow Labs cause banding of phaeomelanin pigment in the hair shafts and as a result, provides the shading effects observed in yellow Labs. Conversely, agouti has no effect on the black/chocolate Lab because Mc1r is always turned on in black/chocolate Labradors, even in the absence of MSH. Therefore, the recessive Agouti alleles will have no effect on the black/chocolate coloration. It is for this reason that one only observes the effects of the Agouti alleles in the yellow Labrador. If the yellow Lab carries the dominant Agouti supressor gene, As, phaeomelanin production will be inhibited, and since eumelanin cannot be made, the dog will appear very pale yellow (nearly white). In contrast, ay or as agouti will inhibit MSH from binding to the receptor and phaeomelanin production will increase.
Some Agouti alleles, such as as, also produce pigmentation patterns that result in more phaeomelanin production on the the dog's back and less phaeomelanin production on the dog's belly. Interestingly, the as allele was not originally proposed by C.C. Little but was later introduced to explain an allele that would encode for the phaeomelanin saddling effect in some breeds that only have tan pigmentation variation on the back and stomach, as opposed to the black and tan saddling effect caused by the at allele in some other breeds. At that point in time that the as was proposed, the prospect of the "loss of function" receptor ("e") had not been explored. However, the aw allele (the "white-bellied" allele) was identified as influencing the expression of Agouti at different concentrations in different locations of the body. Primarily, dogs carrying aw express more agouti protein (for expression of phaeomelanin) on the dorsal surface (the back) and less agouti is produced on the ventral surface (the stomach). The result of aw in a Lab homozygous "e" at the Extension Locus would have the same effect as the proposed as. Therefore, it is possible that as and aw are one and the same allele but expressed differently dependent on whether the breed carries the mutant "E" allele (for eumelanin production only), the "E+" allele (for normal expression/inhibition of eumelanin) or is homozygous for the "e" allele (loss of function receptor/phaeomelanin only). Further modification of phaeomelanin intensity (concentration) will be determined by the products of the C locus which control levels of tyrosinase, an enzyme required in the process of pigment synthesis which preferentially acts upon phaeomelanin. The dominant C allele encodes higher levels of tyrosinase resulting in full intensity of red pigment. Other less dominant alleles encode for less tyrosinase and have the effect of diluting the red pigment to yellow.
Getting Down to Business: The Gene Loci for Canine Coat Color and their Many Alleles
The American Kennel Club acknowledges only three coat colors for the Labrador Retriever breed: black, chocolate, or yellow. The actual color appearance of the dog is said to be its phenotype. Simplistically, however, there are at least nine combinations of alleles just to determine which of the three colors the Lab will appear. This combination of genes is referred to as the genotype.
In the canine, geneticists studying coat color have proposed that there are eleven gene loci controlling coloration. Additionally, each of these loci have multiple alleles. Many Lab breeders and enthusiasts are familiar with the standard method for indicating a Lab's genotype by use of the B/b, E/e nomenclature (refer to A Guide To Coat Color Inheritance In the Labrador Retriever). This standard form indicates only the gene loci responsible for producing the black or chocolate coloration (the B locus) and the yellow coloration (the E locus). For the most part, this system is adequate to predict the expression of simple color traits in offspring produced by two Labs. However, when one wishes to understand more about variations in color intensity or shading within yellows, or the reason for other coat phenotypes that may occur less commonly in Labs, analysis of the other gene loci are required. These other gene loci are the sites for producing modifiers of coat color; that is, their protein products may alter the size and synthesis of melanin (modifiers) or influence the distribution of melanocytes (distributors) to bring about color patterns.
The following table indicates the eleven gene loci and their respective alleles currently accepted as influencing coat color in canines and whether or not they influence phenotypes of the Lab.
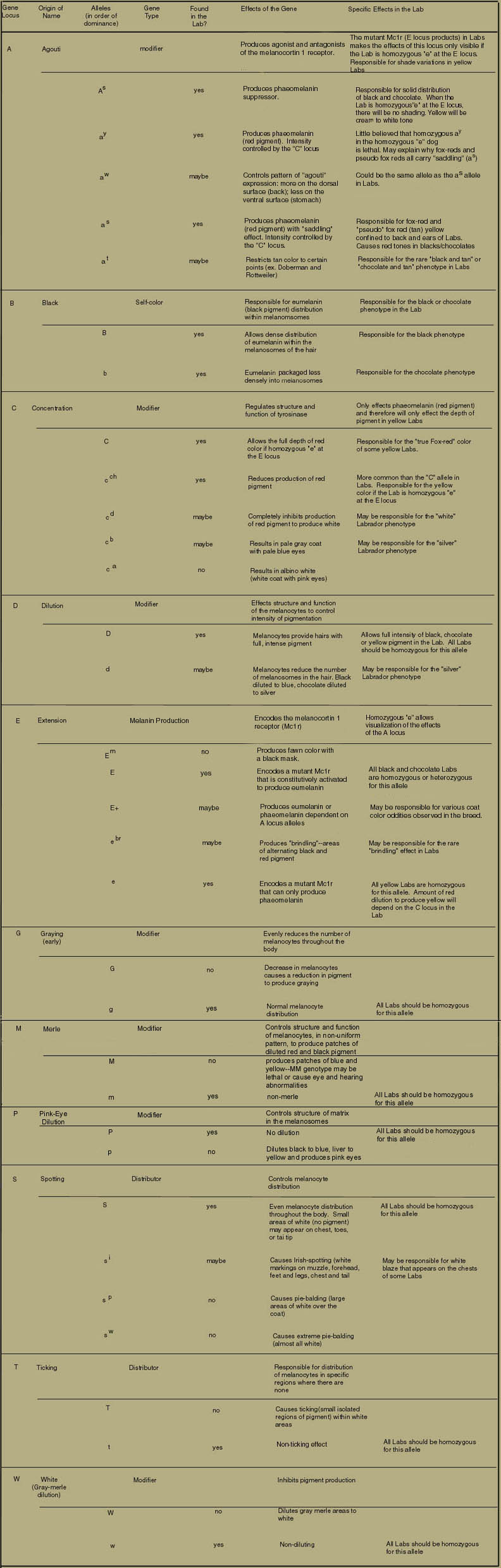
The Yellow Labrador Retriever: A Model for Discerning Effects of Gene Loci other than "B"
The E locus and its "e" allele encodes the mutant receptor that can only produce phaeomelanin (red pigment as observed in the Irish Setter). The product of the A locus and the C locus will determine the location and extent to which this red pigment is diluted to yellow. Therefore, although the "e" allele provides the basis by which the yellow color is apparent, the actual yellow appearance is dependent on the alleles present at both the A locus and the C locus.
Breeders of "true fox reds" will quickly point out that some yellow Labs professed to be "fox-red" are really more dark tan than red and are therefore, not "true fox-reds". The difference in concentration of red color (determined by the "ay" or "as" allele of the A locus) is dependent upon the alleles at the C locus. The "C" allele allows for full expression and intensity of red tones, while the "cch" allele will dilute the red to a clear tan color.
Therefore, the genotype of each color variation is:
ay_B_ C_ee = True Fox-Red
as_B_C_ee * = True Fox-Red with Saddling**
ay_B_ cch _ee = Pseudo Fox-Red
as_B_cch_ee = Pseudo Fox-Red with Saddling**
* the underline denotes that the gene locus may be homozygous or heterozygous with a
less dominant allele present at the partner-chromosome gene locus
** Labs with this genotype demonstrate the red coloring localized to certain areas of the
body.
The "as" allele produces the "saddling effects" seen in
many yellows in which there appears darker yellow pigmentation on the back,
ears, legs, etc. compared to areas of light yellow on the shoulders, neck, and underside.
The "as" allele also increases intensity of phaeomelanin, but
restricts its production to the former mentioned areas on the Lab.
The observation that there appears to be no solid fox-red or solid "pseudo" fox-red Labs may be explained by Little's hypothesis that the combination of an "ay" in a homozygous "e" (yellow) dog is lethal. If Little's hypothesis is correct, then this would mean that all fox-red or "pseudo" fox-red Labs must be: as_B_C_ee or as_B_cch_ee, respectively.
Medium to light shades of Yellow
Medium yellow is probably the most common yellow coloration observed in Labs. The ranges in the shades of the yellow coloration, however, can be quite extensive. Medium yellows, listed from darkest to lightest, are produced by the following genotype combinations:
AsasB_ C_ee
AsasB_ cch_ee
AsAsB_ C_ee
AsAsB_ cch_ee
Those yellows heterozygous "As" "as" will produce less phaeomelanin because the As allele encodes a phaeomelanin suppressor. However, the alleles of the A locus are incompletely dominant, so some phaeomelanin will be produced because of the as allele. The phaeomelanin intensity will be further controlled by the "C" locus, hence the intensity of the yellow may be stronger in these heterozygotes carrying the "C" allele and lighter in those carrying the "cch". The homozygous "As" Labs do not produce red pigment at all because the suppressor will completely block phaeomelanin production by the mutant Mc1r encoded by the homozygous "e" allele. As a result, the coat will appear a cream color and will appear almost white if the "cch" allele is the most dominant allele at the C locus. Additionally, Labs homozygous "As" will show an even distribution of yellow color devoid of shading.
Although the "white" color is considered by most Lab breeders to be a very light shade of yellow, this color may be quite distinct from the yellow shade that may represent cream-colored yellows represented as genotype [As_B_ cch_ee] above. In fact, the "white" color may be represented by another allele that may be found at the C locus. The "cd" allele is responsible for producing white hair in other breeds of dogs, like the West Highland White Terrier, while allowing full expression of dark nose and eye pigment. Though this white color may be distinct from the yellow coloration, it should still be grouped with the other yellow variations since its expression is also controlled by both the E and C loci.
Geneticists recognize that there are two gene loci that are capable of controlling production of the yellow color in the dog. In Labradors, homozygous "e" at the Extension loci is considered the predominant genotype for producing yellow. However, in some other breeds, homozygous "ay" at the Agouti loci is recognized as being responsible for producing the yellow (tan/sable) color in dogs with the wild-type (E+) Mc1r, such as in the Basenji, Collie, Dachshund, etc.
The yellow (buff) color of the Cocker Spaniel was once believed only to be determined by homozygous "e" just as in the Labrador. Interestingly, however, upon occasion when two yellow Cocker Spaniels are bred, a black puppy will be produced. This observation was first made by Clarence Little in 1957 and later confirmed by Burns and Fraser in 1966. Because these test breedings were controlled studies, the possibility of mismating as an explanation was ruled out and a new hypothesis was postulated: There are two kinds of "yellow" Cocker Spaniels, an AsAsB_ee Cocker Spaniel that is usually buff-colored and an ayayB_E+E+ Cocker Spaniel that is usually sable-colored. When these genotypes are crossed, one possible resulting genotype of the offspring will be AsayB_E+e: a black Cocker Spaniel!
This scenario may not be limited to the Cocker Spaniel breed. Occasionally, black puppies are produced from yellow Lab X yellow Lab crosses. Some Lab breeders immediately cry "mismating", however, mismating is clearly not the only explanation since many times mismating is ruled-out by virtue of circumstance (ie. the bitch was exposed only to the intended stud and there was no opportunity for breeding to occur with any other male). In addition to what has been observed for the Cocker Spaniel, there may be additional indications supporting this theory. One author has suggested that the way to distinguish between a homozygous "e" yellow and a homozygous "ay" yellow is to examine the whiskers: If the whiskers are cream or straw colored the dog is homozygous "e", if the whiskers are black then the dog is homozygous "ay" (refer to: "Canine Color Genetics" by Sue Ann Bowling).
It is possible that like the Cocker Spaniel breed, the Labrador has two genotypic "kinds" of yellow dog: one that is homozygous "e" (more common) and one that is homozygous "ay" (less common) with the wild-type Mc1r (E+). As such, crossing these two different genotypic types of yellow Lab could produce an occasional black puppy from two yellow parents. This may also explain why occasionally a black puppy is whelped in litters from a chocolate to chocolate cross. It is also conceivable that some of these ayay Labs, especially if they are homozygous "C" at the C locus may appear to be chocolates rather than yellows (albeit with a more red-tone than a brown tone). As such, crossing of one of these chocolate-appearing yellow Labs with a true chococlate would produce AsayBbEE+: a black Lab.
While on the Subject of Yellow Labs…
Why is it that the yellow Labs' ears are always darker than their bodies, even when they have no shading on their bodies? And what causes the noses of yellow Labs to fade during winter months?
The answers to both these questions can be found by examining the tyrosinase enzymes responsible for producing melanin from tyrosine. Some forms of these enzymes are temperature-unstable mutants that only produce melanins under ideal temperature conditions. Some tyrosinase enzymes work more efficiently in colder temperatures. Extremities, like the ears, are usually a cooler temperature than other parts of the body and as a result, the tyrosinase is able to produce more pigment in this region.
Conversely, the tyrosinase enzyme responsible for producing the dark nose pigment in yellows is unstable at low temperatures. Under conditions of low temperature, the tyrosinase enzyme stops driving the chemical reaction, and tyrosine conversion to eumelanin in the skin will occur at a much slower rate. As a result, pigment will fade.
Though certain drugs may also produce pigment fading, this latter cause for reduction in pigment occurs because these drugs will bind to dopa (an early precursor to melanin in the reaction from tyrosine to melanin) and inhibit the further chemical reactions that result in melanin. This condition will also cause fading of pigment in the Lab.
Unique Variations Occurring in Labrador Coat Color
Color oddities have occurred occasionally throughout the breed history of the Labrador Retriever. Such variations on the typical black, chocolate or yellow coloring have included but are not limited to black-and-tan points, brindling, and silver-casting. It is important to recall that during the early and perhaps mid-history of the breed, interbreeding with other breeds occurred. Crossbreeding to breeds that carry the "wild-type" extension allele (E+) as well as the possibility of a spontaneous mutation resulting in a "gain of function" (also denoted E+) of the Mc1 receptor encoded by the recessive "e" allele, may be possible explanations for color oddities occurring in the breed.
The E+ allele (whether wild-type or "gain of function") would encode a normal, functional Mc1 receptor that would be under greater influence of the alleles at the Agouti (A) locus. Unlike the E allele mutant found normally in Labs that does not require activation by MSH to produce eumelanin (and therefore is always "turned-on" producing black or brown pigment), the Mc1 receptor encoded by E+ would be dependent upon MSH for production of eumelanin (black/brown pigment). Homozygous E+ would allow the effects of the Agouti locus that would otherwise not be seen in a Lab carrying the typical mutant "E" allele (in the typical Lab, effects of Agouti are only seen if the dog is homozygous "e"). Therefore, a Lab that is homozygous E+ would also have to carry As to appear solid black (or chocolate dependent on the allele at the B locus). If a recessive allele (ay, at or as) were the most dominant allele at the A locus, then the Lab would appear either red/yellow (ay or as) or black-and-tan (at). Other Agouti alleles such as aw (which is attributed to producing silver in some breeds) may also be observed.
Early breeding records indicate that a Labrador puppy with tan points on the ears, muzzle, and above the eyes (as found in the Doberman and Rottweiler) would occasionally be whelped to pure-bred Labrador parents. Breeders attributed this to previous interbreeding of Labradors with Gordon Setters during the early history of the breed. Because this trait was considered undesirable as a characteristic of the breed, breeders chose not to breed individuals expressing the trait in hopes of reducing frequency of its expression in future offspring.
Today, it is recognized that tan points are controlled by the "at" allele of the A locus and that it is recessive to most of the alleles found at the A locus of Labs. Because this allele is recessive, it may be passed through many generations before a breeder is aware that the allele is present. In order for the allele to be expressed, a carrier would have to be bred to another carrier of this same allele and both parents would have to be carriers of the wild-type (E+) Mc1 receptor. This explains the low frequency of expression of this trait in the current Labrador population.
Brindling Effects And "Mosaics" in Labs
Brindling describes alternating expression of black and red color in the hair throughout the coat. There are several possible causes for this fault that occasionally appears in Labs. One cause may be attributed to the "ebr" allele that controls brindling in many other breeds of dogs. For expression of this trait, both sire and dam would have to carry the mutant "ebr" allele, which is recessive to the "E" allele, but more dominant than the mutant "e" allele for yellow.
Alternatively, brindling in Labs may be the result of what geneticists call a mosaic. A mosaic indicates differences in the somatic tissue of heterozygotes that come about during mitotic division of somatic cells (recall from above that somatic cells are those that make-up the body). There are two possible ways by which an individual may become a mosaic. The first is called chromosome nondisjunction by which during division into daughter cells, one of the chromosomes fails to separate from its duplicated chromosome. As a result, one daughter cell receives an extra chromosome and the other receives an unpartnered-chromosome.
The second way that a mosaic may be produced is called chromosome loss by which the chromosome containing the dominant allele gets left behind when the daughter cell's nucleus reconstitutes.
In either situation described above, the daughter cells of these altered somatic cells will contain the same alterations. As a result, one will observe a mosaic or brindled pattern of normal color mixed with color produced by the altered somatic cells. This condition has been reported in a Lab showing mosaic black and yellow coat color. When this Lab was bred to other Labs of normal coat colors of black, chocolate, or yellow, it was determined that the variation in color was not due to a mutated E locus allele (like the "ebr" allele) because none of the offspring demonstrated this phenotype. Rather, this coat characteristic was attributed to a chromosomal alteration as described above.
Therefore, the brindling phenotype rarely observed in Labs might be the result of a stable allele mutation (such as the "ebr" allele), or a random somatic chromosome mutation involving the E or B loci. To view an example of a mosaic occurring as a result of a random somatic chromosome mutation involving the E locus in the Labrador Retriever click here.
The silver coat color in Labradors has gained much attention recently and is a very controversial topic (see The Labrador Coat Color Controversy: Do Silver Labs Really Exist?). Reasons for the controversy stem from the lack of information available to trace the origins of this color in the breed as well as the fact that the AKC standard for the Labrador breed does not acknowledge silver as an acceptable color for a Lab. Some breed enthusiasts consider the silver coloration to be a sign of impurity of the bloodline, however, what geneticists have come to understand of recessive alleles is that they may be passed through many generations going undetected, such as the allele for tan points discussed above.
The range observed in silver coloration suggests that silver occurs through a modifying gene. There have been several possible outcomes observed for the silver Lab:
Black Lab + silver modifier = charcoal gray coat with a "sparkly"-like appearance. Nose: dark gray; Eyes: dark to light gray
Chocolate Lab + silver modifier = "mousy"-brown gray coat. Nose: same as coat; Eyes: yellow to gray-yellow
Yellow Lab + silver modifier = platinum to pale silver (yellow with gray casting). Ears: gray (instead of red-toned); Nose: dark to pale gray; Eyes: dark to pale gray.
There are several possible explanations for the silver coat color in Labs. The first explanation would attribute this rare color in the breed to the D locus. Recall that the alleles of the D locus modify the color determined by the B locus. Therefore, if a dog is homozygous or heterozygous for black at the B locus, presence of homozygous recessive "d" at the D locus would dilute the black pigment to appear blue. Alternatively, if a dog is homozygous for chocolate at the B locus, presence of homozygous recessive "d" at the D locus would dilute the chocolate pigment to appear silver. The absence of the corresponding "blue" phenotype in the breed, however, would seem to argue against this explanation.
Another explanation for silver coat color in Labs would attribute this color to the C locus. There is an allele mutant at the C locus that has been determined to cause silver coat color and blue eyes in dogs. The "cb" allele is believed to be a type of albinism. Since alleles at the C locus influence red pigment only, effects of the "cb" allele should only be observed in dogs homozygous "e" at the E locus. Therefore, a silver Lab would not only have to receive the yellow allele from both parents, but also receive the silver allele from both parents (which is recessive to the common "cch" allele). This allele would explain the silver-toned modification of coat observed in yellow Labs in the presence of the recessive "e" allele, however it would not explain the eumelanin modification in the black or chocolate-based silvers (since the C locus alleles primarily dilute phaeomelanin).
Likewise, the possibility of a "partial loss of function" mutation that may have occurred in the dominant "E" allele resulting in muted tones of eumelanin would not explain the modification of phaeomelanin (yellow).
An alternative explanation for explaining the modification of both eumelanin and phaeomelanin again returns to the wild-type/gain-of-function "E+" allele that encodes for a normal functioning Mc1 receptor. If this allele either occurred as a spontaneous mutation or was introduced into the breed through interbreeding, this might explain the modification occurring in all three colors, particularly when one considers the following:
When one traces the pedigrees of some silver Labs, one finds a history of other color oddities occurring in some related bloodlines to the silver Labs. Occurrences of "black-casting" in chocolates, muted chocolate coloration ("card-board box" coloring), as well as the occasional occurrence of black puppies being whelped from two chocolate parents suggests that these "chocolates" were probably not chocolate at all but rather E+ yellows. As such, it is conceivable that the Agouti alleles could produce an intense red pigment resulting in deep red (interpreted as chocolate especially in the absence of "saddling" modifiers) or diluted, muted red (card board box color) due to further modification by the alleles of the C locus). In black Labs, an ayayEE+ geneotype could produce a muted black color (because of the presence of both receptor types) especially if the alleles at the C locus were cch, thus resulting in a deep charcoal, silvery coat appearance. This suggests a possible role of E+ for the silver coloration as well as for a multitude of other coat color variants that occasionally occur in the breed.
There are several conditions that can produce white hair in Labs. Some of these conditions are determined by color genes and others may be caused by environmental factors that effect melanin production.
To analyze the reason why some black Labs have only a few, not-easily-seen, white hairs on their chests while others have small white spots, it is best to first picture that all Labs are white--the condition of having no melanin production. The gene loci for color control both the color of the pigment as well as the distribution of melanocytes throughout the body of the Lab. Therefore, in a black Lab, although color is determined by alleles at the B locus, alleles at the A and E loci determine even distribution of the color over the entire surface of the coat. Labs that carry an allele other than "As" at the A locus, have a greater likelihood of expressing more white hairs than those Labs that do carry "As". Therefore, although all Labs should be homozygous for the S allele at the S gene locus, some may still express white hairs on the chest, bottom of the feet and under the arms and groin areas.
Unlike the even distribution of graying observed in some breeds of dogs as they get older, graying in Labs, particularly noticeable in blacks and chocolates, occurs in distinct areas such as the muzzle and paws. Graying may occur as a result of certain genes or as a normal process of aging. In regard to genetic causes, the dominant "G" allele is responsible for causing reduction in the number of melanocytes so that melanin production is diminished. However, in the breeds carrying the graying allele, this effect on pigment is an early event, therefore, it is unlikely that the graying observed in older Labs is a result of this allele.
Alternatively, the process of aging is also associated with loss of melanocytes and reduction in pigment production. Melanocytes in the muzzle and paws of the Lab may be more prone to the aging process and thus gray more quickly than other locations of the body. Additionally, the reduction in blood circulation and resulting cooling of extremities in geriatric Labs may reduce production of melanin under the control of cold-temperature-unstable tyrosinase enzymes as explained above.
Other potential causes for white and gray hairs include tissue injury that may destroy the melanocytes in a particular area as well as dietary deficiency of copper, which is required for the production of melanin. The former would appear as localized depigmentation, the latter would appear as an evenly distributed loss of pigmentation.
White "Ring around the Tail" and Two-Toned Appearance in Black Labs
Helen Warwick reported the former condition occurring in Labs in her book "The Complete Labrador Retriever." In one chapter, she describes that some Labs have white at the base of the hair shafts on the tail. This white color is only discernable when one lifts up the hairs to view the base of the hair shaft closest to the skin. Mrs. Warwick made note that she had observed this condition primarily in Labs of English descent.
This condition, which is often not limited to the tail, is actually found frequently in both black and chocolate Labs and most likely indicates that the Lab is heterozygous at the Extension (E) Locus ("Ee") and does not carry the As allele at the A Locus. These Labs also can be observed as having red tones in their coat (occasionally causing a two-toned appearance especially obvious during shedding season). This two-toned appearance is not attributed to chocolate undertones, as some breeders may believe, but is rather due to the production of the red pigment, phaeomelanin, in "e" (yellow) carriers that also carry "ay-" or "as-".As a result, both Mc1 receptor types will be present in the dog: one that continuously makes eumelanin and one that will only make phaeomelanin. The receptor encoded by "E" for production of black/brown is more efficient, however, some phaeomelanin will be produced by the "e" receptor. Since the recessive Agouti alleles control synthesis of phaeomelanin during particular times of hair growth (with phaeomelanin produced only during the mid to late portion of hair growth), phaeomelanin will only be seen at the base of the hair shaft. If these Labs also carry the "cch" as the dominant allele at the C locus, the red pigment will be diluted allowing the base of the hair shaft to appear very light in color.
To date, much of the knowledge pertaining to coat color inheritance is very limited with most of the information either derived from small canine studies or extrapolated from other species. Therefore, much of the data is still open to interpretation. In light of this, this article presents the current consensus for inheritance of coat color based on the previous findings and interpretations of those in the field. It is anticipated, however, that our knowledge of canine genetics will be greatly enhanced over the next several years as scientists conducting the "Canine Genome Project" draw closer to their goal of mapping specific genes to their chromosomes in the canine.

References
Newton JM, et al. Melanocortin 1 receptor variation in the domestic dog. Mammalian Genome, 11:24-30, 2000.
Everts RE, Rothuizen J, van Oost BA. Identification of a premature stop codon in the melanocyte-stimulating hormone receptor gene (MC1R) in Labrador and Golden retrievers with yellow coat colour. Anim Genet, 31:194-9, 2000.
Lu D, et al. Agouti protein is an antagonist of the melanocyte-stimulating-hormone receptor. Nature, 371:799-802, 1994.
Vage DI, et al. Molecular and pharmacological characterization of dominant black coat color in sheep. Mammalian Genome, 10:39-43, 1999.
Vrieling H, et al. Differences in dorsal and ventral pigmentation result from regional expression of the mouse agouti gene. Proc.Natl.Acad.Sci. USA, 91:5667-5671, 1994.
Cornell University Animal Health Newsletter, "Coat Color Inheritance," Volume 14, Number 11, January 1997.
Sponenberg DP and Bigelow BJ. An extension locus mosaic Labrador retriever dog. Hered., 78:406, 1986.
Suzuki, D.T., Griffiths, A.J.F, and Lewontin,
R.C. (eds.), An Introduction to Genetic Analysis,San Francisco, W.H. Freeman and Co.,
1981.
Fitzpatrick, T.B., Szabo, G., Wick, M.M., Biochemistry and Physiology of Melanin
Pigmentation, 1981.
Helen Warwick, The New Complete Labrador Retriever, New York, Howell Book House, Inc.,
1986.
Helen Warwick, The Complete Labrador Retriever, New York, Howell Book House, Inc.,1965.
Leon F. Whitney, DVM, How To Breed Dogs, New York, Howell Book House, Inc., 1972.
Little, C.C., The Inheritance of Coat Color in Dogs. Comstock Publishing Associates, Ithaca, 1957.
Willis, M.B. Genetics of the Dog. MacMillan Publishing Co., New York, 1989.
Copyright © 1999. Pamela A. Davol. All rights reserved. Copyright & disclaimer.
Back To Main Menu